본격!! 식물 생태와
생물 상호작용
한반도 식물의 분포 및
종분화 이해의 새로운 관점
생물지리구역의 중요성
생물상에 기초하여 생물지리구와 그들의 공간 범위 및 양상을 이해하는 것은 시간에 따른 생물상 분화를 이해하고 생물다양성의 공간적 양상을 창출한 생태진화적 원동력을 더 깊게 이해하는 필수 작업이다 (Ricklefs, 2004). 특히 생물다양성의 공간 구조를 이해하는 중요한 단계로서 그것의 공간 범위의 추출은 생물지리학, 생태학, 지학 및 보전생태학에서 기초 및 응용 연구의 바탕이 되어 왔다 (Brum et al., 2017; Graham and Hijmans, 2006; Ibanez-Erquiaga et al., 2018; Kreft and Jetz, 2010; Lenormand et al., 2019; Olson et al., 2001; Ricklefs, 2004; Sun et al., 2008).
생물상구역화 (bioregionalization)는 자료의 유형과 규모에 따라 종 및 기후자료가 포함된 생물군계 (biome), 특정한 분류학적 그룹의 분포를 대상으로 작성된 생물지리지역 (biogeographical region) (Linder et al., 2012), 전체 식물종을 대상으로 한 식물상 구역 (floristic zone) 등으로 구분된다 (Born et al., 2006; Gonzalez-Orozco et al., 2014). 최근까지 식물을 통한 지리학적 공간 설정을 위해 많은 제안들이 이루어졌다. 그러나 제한된 자료와 작성의 편의성, 그리고 전문가 견해를 통한 생물학적 지역들이 제안되어 왔었다 (White, 1983). 명확한 기준 (criterion)의 제안은 없었지만 고유 식물종 (endemic plant)을 활용한 공간 설정 (Takhtajan, 1986), 그리고 식물상 조사 자료 (floristic checklist)의 축적과 공간 통계 기법의 발달에 따른 식물구의 제안들이 존재해 왔다 (McLaughlin, 1989).
지역 생물다양성 양상의 이해를 위해 필요한 정밀한 지도화 작업과 결과 생산에는 여전히 어려움이 존재하는데, 그것은 특히 식물상 조성 조사의 충실성과 신뢰성 확보의 어려움, 그리고 식물상은 환경 경사 (개발 또는 기후) 및 그의 복잡성과 매우 밀접하게 연결되어 있기 때문이다 (Gonzalez-Orozco et al., 2014). 이러한 도전들과 함께, 생물지리학적 지역화의 중요한 기초 단계로서 충실하고 신뢰성 높은 자료를 통한 생물 공간의 추출은 새로운 기준 (metrics)과 관점을 제공할 것이다 (Lenormand et al., 2019).
한반도에서 첫 생물지리학적 공간화는 백 여 년 전 일제강점기에 식물상과 식생 특성에 관한 설명을 편리하게 하기 위해 북부, 중부 및 남부 구분이 제안되었다 (Nakai, 1919). 이후 Nakai (1919)의 구분을 재분할하며 세분된 식물구들이 제안되었고 (Lee and Yim, 1978), 식생-기후 관점에서 유사-식물지리구의 공간이 제안된 바 있다 (Yim and Kira, 1975). 주로 전문가 경험 또는 편리함이 반영된 기존의 한반도 식물구 (유사-식물지리구 포함)들은 모두 띠 또는 균질한 면 모양 (위도 및 연평균기온 상관의 관점)으로 제안되었다.
생물지리적 경계의 추출은 영역들 간의 차이를 최대화하면서 영역 내 분류학적 조성의 균질성을 최대화 해야 한다 (Stoddart, 1992). 식물구와 같은 생물 구역의 추출이 직면한 도전이었던, 생물 분포 자료의 양적 축적과 지리정보화, 그리고 대량의 생물 분포 자료를 활용하는 분석 능력의 향상은 정량적이고 정확한 공간 추출을 가능하게 한다 (Linder et al., 2012). 최근의 연구에서 역시 축적된 자료들을 활용하여 분포 정보를 부여하는 방식으로 식물구를 추출하고 있다 (Gonzalez-Orozco et al., 2014). 생물 분포 점 자료의 신뢰성은 그들의 표본이 가장 우수하다. 20세기 후반 위성항법장치 (Global Positioning System, GPS)의 보급과 함께 공간 정보가 기록된 식물 표본들이 축적되고 있으며, 이 분포 점들을 활용한 신뢰성 높고 정확한 식물구 추출에 관한 접근들이 필요하다. 특히, 한반도 남부에서 2000년 이후 진행된 식물 표본 수집과 분포도의 구성은 신뢰성을 갖춘 대규모 자료로 평가할 수 있다
식물상 조합에 기초한 새로운 구역들
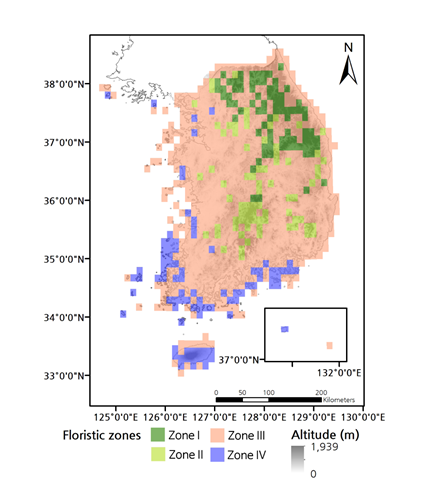
2003년부터 2015년까지 수집된 식물 분류군의 표본과 좌표 정보를 바탕으로 2016년 발간된 한반도 남부 관속식물 분포도는 175과 919속 2,954분류군에 대한 309,333 건의 좌표 정보를 담고 있다 (Korea National Arboretum, 2016). 식물 표본의 좌표 정보를 771개 격자의 식물상 조성 (floristic assembly) 자료로 변환하여 추출한 한반도 남부의 식물상 구역은 4개 공간으로 구분하는 것이 제안된다. 내륙 구역 (Zones I~III)들은 한반도 중부의 고해발 지역인 한랭 식물상 구역 (Cold floristic zone), 한반도 남부의 고해발 지역인 상량 식물상 구역 (Cool floristic zone), 그리고 한반도 중부 및 남부의 저지대인 중남부 온난 식물상 구역 (Warm floristic zone)으로, 그리고 제주도 및 울릉도가 포함되는 연안 식물상 구역 (Maritime floristic zone)으로 구분되었다.
한반도 남부에서 175과 919속 2,954분류군 309,333 건의 좌표 정보를 토대로 구분한 새로운 식물상 조합의 구역은 4개로 구분된다 (왼편). 현재 활용되고 있는 한반도 식물상 구역 (식물구계)은 일제강점기 일본 식물학자 나까이 다케노신이 한반도 북부, 중부 및 남부와 같이, 생물지리학적 관점을 투영하기 보다는 단지 식물상 설명의 편의성을 위해 설정한 구분선들이 시초가 되었다.
이후 식물상 자료의 축적과 학자들의 관점이 더해져 구분선들이 추가되었고, 현재 활용되고 있는 식물구계도가 되었다 (오른편의 지도). 현재 한반도 식물구계도 및 파생된 식물 구계학적 특정종은 생물학 및 지리학적 의미 (종분화, 지형 및 기후)는 뚜렷하지 않지만, 1990년대 제정된 환경영향평가법 (환경부)에서 그것을 활용하도록 강제하였기 때문에 꾸준히 활용되고 있다.
추출된 공간들의 양상을 통하여 한반도 남부 지역의 전이적 (온난으로부터 한랭) 식물상 변동 진행과 물리적 및 인간 활동 영향이 확인되었다. 한반도의 역사적 식물상 구역은 크게 기후 요인과 식물 분포 양상을 복합적으로 반영하여 구성되었고, 면에 가까운 띠모양이다 (Lee and Yim, 1978). 새로 제안된 Zone I~III는 역사적 구역들과는 완전히 다른 모습을 보인다. 역사적 중부 및 남부 구역은 상록활엽수 (예, 탱자나무속 Genus Citrus 및 감탕나무속 Genus Ilex)의 분포와 같은 일부 수목 생태형의 분포 양상을 일부 반영하였지만, 산들의 복잡한 묶임도 및 해발고도 효과와 같은 소규모 지역에서 중요한 생물지리학적 요인들의 고려는 제한적이었다. 최근 관련 연구들은 띠모양 보다는, 빙하기 – 간빙기 변동과 같은 기후 및 물리 환경의 변동, 그리고 지형과 같은 다양한 요소들의 역할에 따라, 생물상 조합의 구역 내에 이질적이고 소규모 군반들이 다수 존재하는 공간 분포 양상 (지구 및 지역 규모 모두)을 나타내고 있다 (Kreft and Jetz, 2010; Lenormand et al., 2019; Médail and Diadema, 2009; Silva and Souza, 2018).
한랭 식물상 구역 Zone I은 한반도 중부 내륙의 고해발 산들 (최고 해발 1,708 m Mt. Seoraksan)을 중심으로 추출되었고, 일부 남부 고해발 산들이 포함되었다. 역사적 구역에서는 중부 및 남부 구역의 일부에 속한다.
Zone I은 과거 주빙하 환경에서 남하했던 식물상 요소들이 그것의 후퇴화 더불어 한반도 중부 고해발 산맥에 잔존하고 있는 식물상 구역이다. 또한 동아시아에서 눈잣나무 (Pinus pumila), 눈측백 (Thuja koraiensis) 및 분비나무 분포의 남쪽 경계 지역이다 (Kong et al., 2019; Korea National Arboretum, 2015). 또한 서늘한 기후를 활용한 고랭지 농업 highland agriculture 그리고 임목 생산 (잣나무 및 일본잎갈나무)이 매우 활발한 지역이다. Zone I보다 위도가 높은 공간들이 Zone III으로 추출되는 것은 강한 군사 활동이 오랜 기간 진행된 지역이기 때문이다.
상량 식물상 구역 Zone II는 Zone I과 인접한 산지들, 그리고 대부분 한반도 남부 내륙의 고해발 산 (1,915 m 지리산 및 1,614 m 덕유산)들이 중심이다. 역사적 구역에서는 Zone I과 동일하게 중부 및 남부 식물상 구역의 일부에 속한다. 구상나무 분포의 북쪽 한계, 그리고 동아시아에서 가문비나무 (Picea jezoensis) 분포의 남쪽 한계가 포함된다 (Korea National Arboretum, 2015). Zone I과 동일한 지사적 과정으로서, 주빙하 환경의 영향이 한반도 남부 지역에 잔존하고 있는 공간이다. 또한 보다 따뜻한, 또는 해양성 기후에서 주로 분포하는 수목들 (예, 노각나무 및 털조장나무) 역시 Zone II의 중해발 및 고해발에 지역에 생육하고 있다 (Kim et al., 2014). Zone II는 고랭지 농업은 거의 없으며, 일부 임목 생산 (주로 일본잎갈나무)이 이루어지며, 인접하여 대도시들이 다수 존재한다.
온난 식물상 구역 Zone III은 한반도 남부의 대부분을 차지하는 식물상 조합의 구역이다. 이 구역 내에는 높은 산들과 산맥이 존재하지만 비교적 토지 이용 강도가 높은 도시 및 농업 도시들의 영향이 복합적으로 작용하는 공간으로 판단된다 (참고자료 4). 주목할 사항으로서, Zone III에는 나머지 식물상 구역 (Zone I, II, 및 IV)을 구성하는 식물들이 하나의 섬과 같이 고립되어 잔존하고 있다. Zone III 내부에 섬과 같은 이질적 식물상 공간들은 지사적 수준에서 식물 다양성 분포 양상의 변화에 따른 하나의 잔존 공간들로서, 지역 식물상 조성의 이질성 형성에 중요하다 (Laliberté et al., 2014; Zobel, 1997). 한반도의 핵심 산이 많은 지형에 의한 생물다양성 피난처 기능은 최근 강조되고 있다 (Chung et al., 2018). 이러한 이질적이고 소형 공간들의 생물다양성 보존 기능 (예, 피난처), 그리고 장기적 변화에 대한 심층 연구가 뒤따라야 한다.
연안 식물상 구역 Zone IV는 세분화되어 활용되었던 역사적인 한반도 남해 연안, 제주도 및 울릉도 식물상 구역들을 통합하며, 특히 해당 구역에만 출현하는 특정 분류군에서 난초과, 국화과, 장미과의 구성 비율이 높은 공간이다.
Zone IV는 일부 내륙 지점들을 포함하지만, 한반도 연안 지역에서 생물다양성의 중요 지역들로서 대양섬 울릉도 (Chang and Gil, 2014; Choi et al., 2019; Holman et al., 2017), 그리고
상록활엽수 (예, Castanopsis sieboldii)의 온난대에서 극지 수목 (예, Diapensia lapponica var. obovate)의 한랭대에 이르는 크고 높은 해발의 한라산 (1,950 m)이 있는 제주도와 같은 여러 섬들로 주로 구성되어 있다. 중요 사항으로서, Zone IV는 한반도 식물상 구역에서 상대적으로 높은 특정 분류군의 다양성, 그리고 난초과의 핵심 분포 공간으로서 보전 및 진화적 관점에서 중요한 위치를 차지한다. 화산섬인 울릉도 및 제주도가 포함되어 다른 공간들에 비해 모재 (volcanic ash), 지형 (lava terrace)을 비롯한 물리적 조건들이 크게 다르다. 제주도의 북부는 남부에 비해 오랜 기간 농업과 행정 중심지의 기능을 수행하였으며, 이것은 식물상 차이의 배경일 수 있다.
<참고문헌>
Born, J., Linder, H.P. and Desmet, P., 2006. The Greater Cape Floristic Region. Journal of Biogeography, 34(1): 147-162.
Brum, F.T. et al., 2017. Global priorities for conservation across multiple dimensions of mammalian diversity. Proc Natl Acad Sci U S A, 114(29): 7641-7646.
Chang, C.-S. and Gil, H.Y., 2014. Sorbus ulleungensis, a New Endemic Species on Ulleung Island, Korea. Harvard Papers in Botany, 19(2): 247-255.
Choi, H.-J., Yang, S., Yang, J.-C. and Friesen, N., 2019. Allium ulleungense (Amaryllidaceae), a new species endemic to Ulleungdo Island, Korea. Korean J. Pl. Taxon, 49(4): 294-299.
Chung, M.Y. et al., 2018. The Korean Baekdudaegan Mountains: A Glacial Refugium and a Biodiversity Hotspot That Needs to Be Conserved. Front Genet, 9: 489.
Gonzalez-Orozco, C.E. et al., 2014. Quantifying phytogeographical regions of Australia using geospatial turnover in species composition. PLoS One, 9(3): e92558.
Graham, C.H. and Hijmans, R.J., 2006. A comparison of methods for mapping species ranges and species richness. Global Ecology and Biogeography, 0(0): 060831043455001-???
Holman, G. et al., 2017. A New Species and Introgression in Eastern Asian Hemlocks (Pinaceae: Tsuga). Systematic Botany, 42(4): 733-746.
Ibanez-Erquiaga, B., Pacheco, A.S., Rivadeneira, M.M. and Tejada, C.L., 2018. Biogeographical zonation of rocky intertidal communities along the coast of Peru (3.5-13.5 degrees S Southeast Pacific). PLoS One, 13(11): e0208244.
Kim, J.S., Chung, J.M., Kim, S.Y., Kim, J.H. and Lee, B.Y., 2014. Phytogeographic study on the Holocene hypsithermal relict plant populations in the Korean peninusula. Korean Journal of Plant Taxonomy, 44(3): 208-221.
Kong, W.S., Kim, H., Kim, D., Lee, C.H. and Shin, H.T., 2019. Natural history and climate change of dwarf relic shrubs. Advan Publishing Inc., Seoul.
Korea National Arboretum, 2015. Conifers. Ecology of woody plants in South Korea, I. Sumeungil, Seoul.
Korea National Arboretum, 2016. Distribution maps of vascular plants in Korea. Korea National Arboretum, Pocheon.
Kreft, H. and Jetz, W., 2010. A framework for delineating biogeographical regions based on species distributions. Journal of Biogeography, 37(11): 2029-2053.
Laliberté, E., Zemunik, G. and Turner, B.L., 2014. Environmental filtering explains variation in plant diversity along resource gradients. Science, 345(6204): 1602-1605.
Lee, W.C. and Yim, Y.J., 1978. Studies on the distribution of vascular plants in the Korean peninsula. Korean Journal of Plant Taxonomy, 8(Appendix): 1-33.
Lenormand, M. et al., 2019. Biogeographical network analysis of plant species distribution in the Mediterranean region. Ecol Evol, 9(1): 237-250.
Linder, H.P. et al., 2012. The partitioning of Africa: statistically defined biogeographical regions in sub-Saharan Africa. Journal of Biogeography, 39(7): 1189-1205.
McLaughlin, S.P., 1989. Natural floristic areas of the western United States. Journal of Biogeography, 16: 239-248.
Médail, F. and Diadema, K., 2009. Glacial refugia influence plant diversity patterns in the Mediterranean Basin. Journal of Biogeography, 36(7): 1333-1345.
Nakai, T., 1919. Report on the vegetation of the Island Ooryongto or dagelet Island, Corea. In: T.g.o. Chosen (Editor), Seoul, Korea.
Olson, D.M. et al., 2001. Terrestrial Ecoregions of the World: A New Map of Life on Earth. BioScience, 51(11): 933-938.
Ricklefs, R.E., 2004. A comprehensive framework for global patterns in biodiversity. Ecology Letters, 7(1): 1-15.
Silva, A.C. and Souza, A.F., 2018. Aridity drives plant biogeographical sub regions in the Caatinga, the largest tropical dry forest and woodland block in South America. PLoS One, 13(4): e0196130.
Stoddart, D.R., 1992. Biogeography of the Tropical Pacific. Pacific Science, 46(2): 276-293.
Sun, Y., Yan, X. and Xie, D., 2008. A new method of vegetation–climate classification in China. International Journal of Climatology, 28(9): 1163-1173.
Takhtajan, A., 1986. Floristic regions of the world. California Press, Berkeley.
White, F., 1983. The vegetation of Africa. UNESCO, Paris.
Yim, Y.J. and Kira, T., 1975. Distribution of forest vegetation and climate in the Korean penisula. I. Distribution of some indices of thermal climate. Japanese Journal of Ecology, 25(2): 77-88.
Zobel, M., 1997. The relative of species pools in determining plant species richness: an alternative explanation of species coexistence? Trends in ecology & evolution, 12(7): 266-269.
식물생태
임업연구사 조용찬 박사후연구원 정성희