생태 과학 돋보기
기후변화와 생물종의 분포 변화:
래포포트 (Rapoport) 규칙의 이해
변화하는 환경과 생물의 반응
1850년 이후 최근까지 진행된 0.8 ~ 1 °C의 지구 평균기온의 상승은 이미 동물과 식물의 생물계절 현상, 분포 범위 이동 및 군집 구조에 상당한 변화를 가져왔다. 지구온난화로 표현되는 이 현상이 초래한 온대 지역 생물의 뚜렷한 변화는 조류와 양서류의 조기 번식, 철새와 나비의 조기 출현, 봄철 식물의 조기 개화 같은 생물계절학적 반응 등을 포함한다. 지리적 범위 이동에서 추운 환경에 높은 내성을 지닌 생물은 유리한 환경조건을 따라 점점 고위도 또는 고지대로 이동하였다. 동시에 열대와 같은 따뜻한 환경에 적응된 종 또한 고위도로 이동하여 이전에는 생존하기 어려웠던 조건인 온대 지역에서까지 나타났다. 복잡한 종들의 반응이 집합적으로 발현되는 군집 수준의 변화는 특히, 지구온난화 이전 조건에서 이미 열 내성 한계에 근접했을 수 있다. 이것은 열대 지방 생물의 멸종과도 관련 있을 뿐만 아니라 다양한 상호작용 속에 종들의 분포 이동과 기존의 군집 특성을 벗어나는 변화 (사망률과 이입률의 변동, 또는 높이 같은 군집 구조)까지도 설명할 수 있는 가정이다.
미래에 지구온난화로 나타나게 될 생물학적 반응에 대한 많은 예측 연구들이 온도와 관련된 생태학적, 생물계절학적, 유전적 및 생리학적 반응에 대해서 이루어지고 있다. 하지만 우리 “과학자들은 여전히 연구하고 있어 상세한 기후변화의 변동성에 대한 예측을 하기 어렵다”는 것이 현실이다. 특히 생물학자들의 주된 관심사는 온도 상승으로 인해 여러 모델이 직면하는 미래를 예상하고 미소진화 경향을 파악하는 것이다. 이 주제는 생물지리학적 지식에 대한 관심을 다시 불러 일으켰다. 이 잠재성 높은 생태지리학적 지식의 시발점은 다윈을 포함한 많은 학자들의 다양한 기록들이 생산된 19세기 이후이다. 생물학자들은 기후대에 걸쳐 식물과 동물의 발생을 지도화하면서 크기, 부속 기관 및 장기의 비율, 색상, 분포 범위 및 생식 전략의 특성을 공유한다고 확신하였다. 그들은 이러한 관찰을 대표적인 7가지의 생태지리학적 생물 법칙 (표 1)으로 요약했으며, 이 중 대부분은 발견자의 이름을 따서 명명되었다 (그림 1). 우리는 이러한 법칙 중 위도 혹은 고도에 따라 생물 분포에 대해 다루며, 최근까지 관련 연구가 비교적 활발한 Rapoport 법칙에 집중해서 다루어 보겠다.
표 1. 생태지리학적 생물 법칙의 요약
| 법칙 |
핵심내용 |
| Bergmann의 규칙 |
추운 환경(고위도)일수록 체형이 커지고 따뜻한 환경(저위도)일수록 체형은 작아진다.
|
| Rapoport의 규칙 |
식물과 동물의 위도(분포)범위는 일반적으로 고위도(추운 환경)보다 저위도(따뜻한 환경)에서 더 좁다.
|
| Gloger의 규칙 |
습하고 따뜻한 환경에 사는 동물은 춥고 건조한 환경에 사는 동물보다 외형색이 더 어둡다.
|
| Allen의 규칙 |
추운 환경에 적응한 동물은 따뜻한 환경에 적응한 동물보다 신체 부속기관(팔 혹은 다리)이 더 짧은 경향이 있다.
|
| Hesse의 규칙 |
추운 환경에 서식하는 동물은 따뜻한 환경에 서식하는 근연종보다 체중에 비해 심장이 더 크다.
|
| Jordan의 규칙 |
어류는 수온과 등 지느러미 또는 척추의 개수와 같은 대칭 특성이 음의 상관관계를 가진다.
|
| Thorson의 규칙 |
저위도의 저서 해양무척추동물은 많은 수의 작은 알을 생산하는 반면, 고위도에서는 수는 적지만
알의 크기가 커지는 경향을 나타낸다.
|
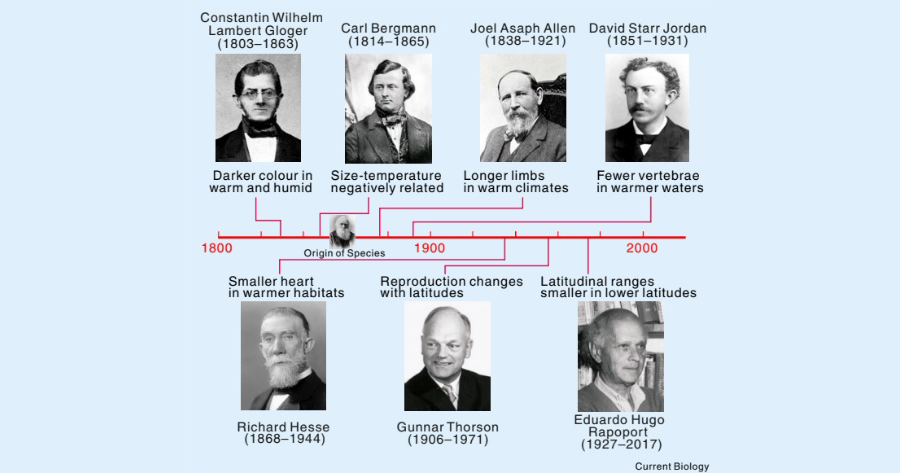
그림 1. 생태지리학적 생물 법칙의 연대표.
1859년 다윈의 ‘종의 기원’ 발간을 기준으로 각각의 법칙이 발표된 날짜에 법칙의 기원인물을 표기함. Tian & Benton. (2020)에서 인용
Rapoport 규칙에 따른 생물 분포
Rapoport 규칙은 식물과 동물의 위도 분포 범위는 일반적으로 고위도보다 저위도에서 더 작다는 것이다. 이 양상은 사지동물, 어류, 해양 연체동물 및 식물의 분포 연구에서 처음 관찰되었다. 이것은 더 나아가 해발고도 차이까지 확장되었고, 낮은 고도보다 높은 고도에서 생물분류군의 분포 범위가 더 넓은 나타났다. 이것은 '기후 변동성 가설'에 의해 설명되었다. 즉, 높은 위도 또는 높은 해발고도와 같은 환경변화의 폭이 큰 서식지 환경에서 분포하는 생물은 더 넓은 환경 내성을 가져야 하며 반대되는 환경 조건에 분포하는 생물보다 더 넓은 범위를 차지할 수 있다는 것이다. 달리 말하자면, 낮은 위도와 고도에서 분포하는 생물은 더 안정적인 서식지 환경에서 좁은 환경 내성 범위를 가진다고 설명할 수 있다.
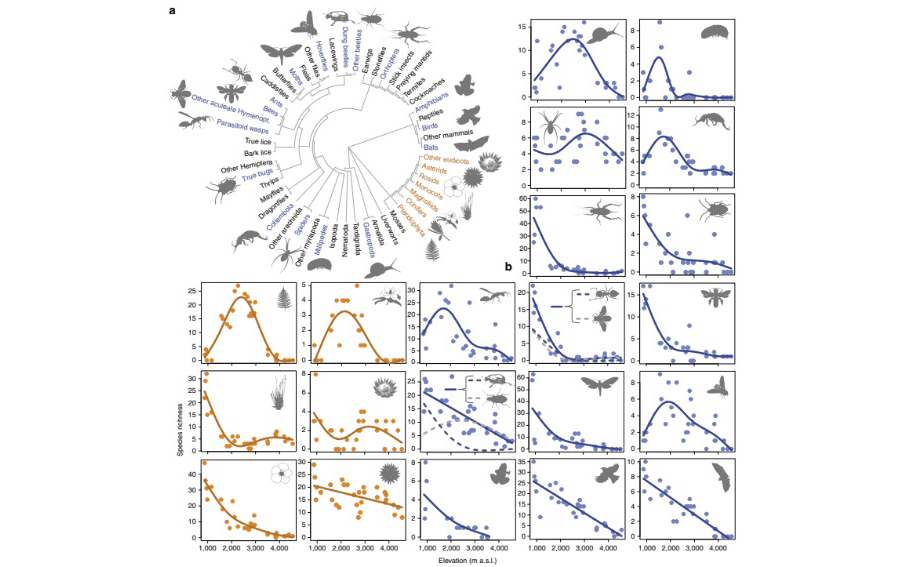
그림 2. 육상 식물 및 동물에서 연구된 분류 집단의 계통 발생 분포;
(a), 그리고 관속식물 (오렌지색) 및 동물 (파란색)의 해발고도에 따른 종풍부도 패턴; (b). Peters, M. K. et al. (2016)에서 인용
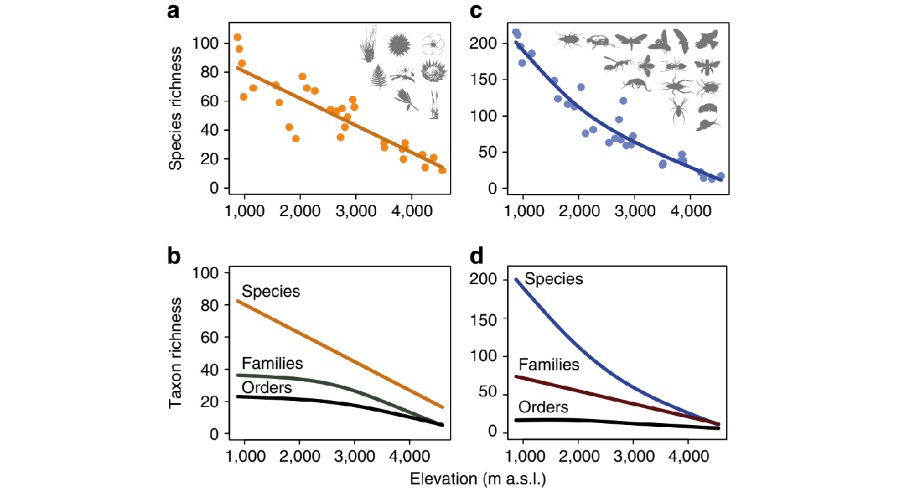
그림 3. 군집 수준에서 식물과 동물의 해발고도에 따른 종 풍부도 양상.
식물과 동물의 종 풍부도는 해발고도에 따라 감소하였으며 (a, c), 마찬가지로 과 단위와 속 단위에서도 감소경향을 보임
(b, d) Peters, M. K. et al. (2016)에서 인용
하지만 이후에 진행된 연구에서 다소 기존의 것과 상충되는 결과가 나타났다. 땅벌 종류의 남쪽 한계 분포는 온난화가 증가함에 따라 줄어들었지만 북쪽 한계분포는 확장되지 않았는데, 이것은 Rapoport 규칙에서 예측한대로 전체 범위 크기가 기온 상승에 따라 축소되는 것을 보여주었다. 그러나 다른 장기 연구에서는 일반적인 지구 온난화에 따라 종의 분포 범위가 높은 위도 또는 높은 고도에서만 발생하여 분포 범위가 증가했다. 고위도 방향으로의 뚜렷한 이동과 범위 확장의 경우 다양성 최고점 역시 높은 위도로 이동하였고, 열대지방 (즉, 저위도)의 생물다양성 최고점이 없다는 것은 열대지방이 급속한 온난화의 결과로 이미 너무 따뜻한 환경으로 변화하였다는 것을 의미한다고 밝히고 있다.
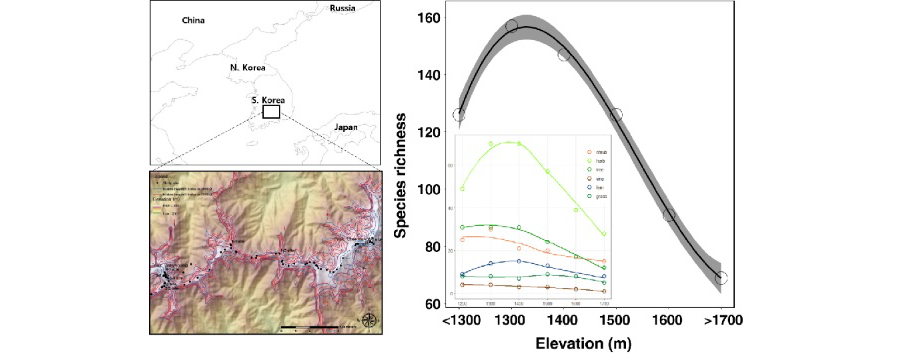
우리 또한 마찬가지로 Rapoport의 법칙에서 말한 분포 범위를 확인하기 위해 우리나라에서 가장 높은 해발고도를 가진 지리산에서 식물 종 풍부도 양상을 살펴보았다. 조사가 이루어진 해발범위의 한계점 (해발 1,300 m부터)이 존재하지만, 일부분이 잘린 역 U자형 특성이 확인되며, 상대적으로 낮은 해발고도 (1300 ~ 1400 m)에서 최고 종 풍부도를 보였다. 식물종의 생활형에 따라 광엽초본 식물종이 전체 종 분포 범위의 변동 양상과 유사하였고, 나머지 생활형의 식물종들은 완만하게 감소하는 양상을 보였다. 우리가 조사한 자료를 분석한 결과로는, 모든 식물종에 대해 Rapoport 규칙에 따르는 경향을 나타냈으나, 개별 생활형에서는 규칙과 다소 상이하거나 약한 경향을 띠었다. 이것은 아마도 우리나라와 같이 복잡한 지형조건이 만들어내는 미세환경의 복잡성, 그리고 생활형 또는 종마다 개별적 성질이 존재하기 때문에 차이를 보였을 것으로 추측된다. 물론 연구의 신뢰성을 위해 다양한 지역에서 동일한 실험 진행이 추가적으로 진행되어야 할 것이다.
그림 4. 지리산에서 확인한 해발고도에 따른 종 풍부도 패턴.
Rapoport 규칙은 통계적 접근이라는 비판과 함께 기본 메커니즘을 가정하여 수많은 논쟁이 이루어졌다. 또한 특정 사례연구는 범위가 위도 또는 고도보다 더 많이 의존하는 요인뿐만 아니라 종마다 적절한 물리적 조건의 지리적 가용성에 따라 어떻게 달라지는지 보여주었다. 변화하는 환경으로 인해 생물의 지리적 범위는 고위도로 이동할 것이며, 이것이 어떻게 될지에 대한 세부사항은 아마도 종마다 다를 것이다. 고위도로의 분포 범위 이동은 범위 증가, 범위 유지, 그리고 범위 감소가 모두 가능할 수 있다. Rapoport 규칙은 분명히 큰 범주에서는 적용되지만 가장자리 범위 내 환경요인 복잡성의 영향으로 강력한 설명 이론이 되지는 않는 것 같다. 기온이 올라감에 따라 기후 변화는 더 다양해질 수 있으며 (예: 극심한 여름과 겨울) 종마다 다른 반응이 나타날 것이기 때문이다.
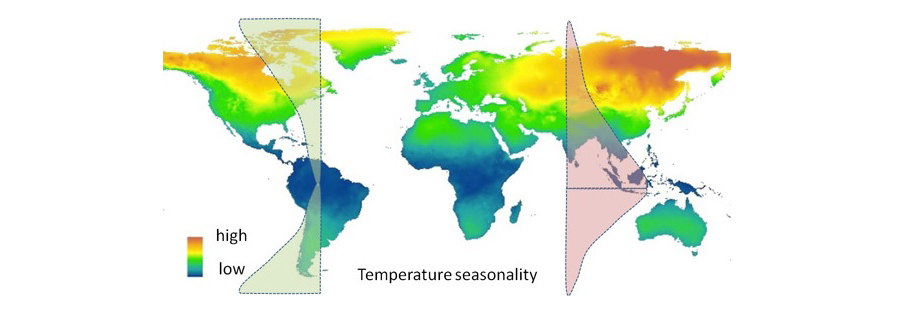
그림 5. 전 세계 온도 계절성 분포에 따른 생물다양성과 분포 범위.
좌측 그래프는 생물의 분포범위를 나타내며, 우측 그래프는 생물 다양성 정도를 설명. http://professor.ufop.br/에서 인용.
변화하는 환경과 미래의 분포 예측
논쟁적이기도 하지만 훌륭한 생산적 규칙으로서, 초기 연구에서 방법론적 약점을 보완하는 추가적인 연구들이 진행되고 있다. 많은 연구들이 식물과 동물이 계통학 (초기 번식), 지리적 분포 (고위도 이동) 및 군집 구조의 변화 측면에서 변화하는 환경에 어떻게 반응하는지 밝혀내고 있다. 이러한 연구들의 일반화는 생물의 생태학적 및 생리학적 취약성에 대한 연구 성과들이 활발하게 생산됨으로써 가능해질 것이다.
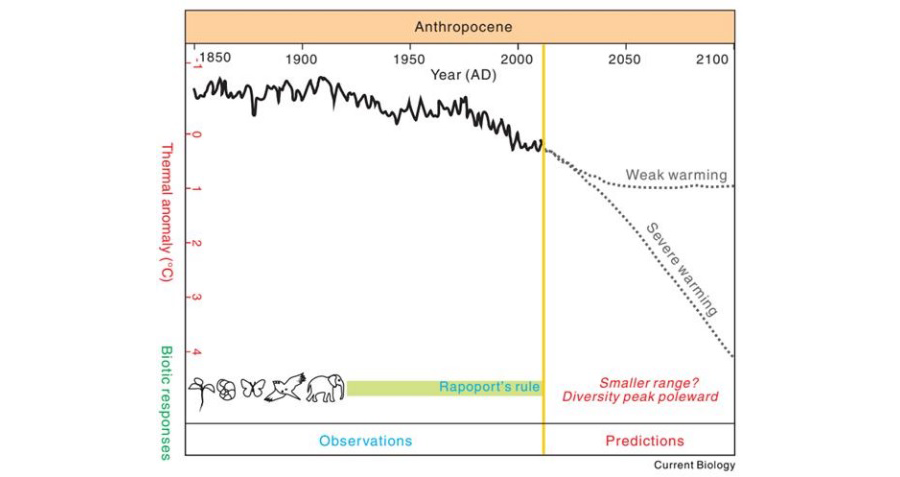
그림 6. 생태지리학적 생물 규칙에 따른 변화하는 환경에 대한 잠재적 생물 반응 예측.
열 이상 곡선은 1850 ~2014년의 육지 및 해양표면 온도변화(실선)와 모델링한 온도변화(점선)로 예측되었으며, IPCC(2014)기준에 따름.
Rapoport의 법칙이 적용되는 생물과 사례연구가 진행된 기간을 표기. Tian & Benton. (2020)에서 인용
변화하는 환경 속에 생물의 지리적 범위는 많은 변화를 나타낼 것으로 예측된다. 또한, 규칙을 따랐던 종의 변화는 '약한 온난화' 및 '심각한 온난화' 같은 시나리오의 성격에 크게 영향을 받을 것이다 (그림 5). Rapoport 규칙 같은 생태지리학적 생물 규칙을 적용한 온난화 관련 생물 변화 예측은 일반적으로 비교적 단순한 규칙의 초기 가정에 비해 결과 해석의 복잡성이 클 수 있다. 특히, 종 보전을 위한 중요한 자료로서 미래의 기후 환경에 대한 생물학적 반응을 예측하는 많은 연구가 있다. 위에서 언급한, 예측된 생물학적 변화가 전 세계 생물 다양성 손실 또는 절멸 같은 치명적인 생물학적 사건과 밀접한 관련이 있다고 말하기 어렵지만, 이러한 예측의 결과는 어떤 종이 생존할 수 있고 어떻게 변할 수 있는지를 이해하는 데 중요하다. 끝으로, 직접적인 관련성을 설명하기까지 합당한 연구들이 진행되어야 하지만 다양한 생물학 법칙 중 많은 부분이 인간에게도 적용될 수 있으므로 지속적인 관심이 필요하다.
<참고자료>
1. Chen, I. C., Hill, J. K., Ohlemüller, R., Roy, D. B., & Thomas, C. D. (2011). Rapid range shifts of species associated with high
levels of climate warming. Science, 333(6045), 1024-1026.
2. Chaudhary, C., Saeedi, H., & Costello, M. J. (2016). Bimodality of latitudinal gradients in marine species richness. Trends in
Ecology & Evolution, 31(9), 670-676.
3. Doak, D. F., & Morris, W. F. (2010). Demographic compensation and tipping points in climate-induced range shifts. Nature,
467(7318), 959-962.
4. Kerr, J. T., Pindar, A., Galpern, P., Packer, L., Potts, S. G., Roberts, S. M., ... & Pantoja, A. (2015). Climate change impacts on
bumblebees converge across continents. Science, 349(6244), 177-180.
5. Parmesan, C. (1996). Climate and species' range. Nature, 382(6594), 765-766.
6. Pintor, A. F., Schwarzkopf, L., & Krockenberger, A. K. (2015). Rapoport's Rule: Do climatic variability gradients shape range
extent?. Ecological Monographs, 85(4), 643-659.
7. Peters, M. K., Hemp, A., Appelhans, T., Behler, C., Classen, A., Detsch, F., ... & Steffan-Dewenter, I. (2016). Predictors of
elevational biodiversity gradients change from single taxa to the multi-taxa community level. Nature communications, 7(1), 1-11.
8. Rohde, K., Heap, M., & Heap, D. (1993). Rapoport's rule does not apply to marine teleosts and cannot explain latitudinal
gradients in species richness. The American Naturalist, 142(1), 1-16.
9. Tian, L., & Benton, M. J. (2020). Predicting biotic responses to future climate warming with classic ecogeographic rules.
Current Biology, 30(13), R744-R749.
10. 김한결. (2021). 한반도 남부 고지대의 식생경관 변화에서 기후변화 영향과 생태적 복원전략. 서울여자대학교 박사학위논문, 서울.
광릉숲보전센터 수목생태연구실
박사후연구원 김한결, 정성희 전문위원 이동혁 임업연구사 조용찬