생태 과학 돋보기
계통발생학적 다양성 (Phylogenetic diversity, PD)의
배경과 간단한 측정 개념
식물의 계통발생학적 다양성은 다른 척도 (α-다양성과 기능다양성)와 마찬가지로 다양한 형질과 생태학적 차이를 통합적으로 측정하고, 군집의 생산성 (Cadotte et al. 2009)과 안정성 (Cadotte et al. 2012)을 포함한 핵심 생태계 기능들과 상호관련을 밝히기 위해 소개되었다 (Srivastava et al. 2012). 계통발생학적 다양성은 생태계의 다기능성 예측에 필요한 정량화 도구로 활용할 수 있으며, 전통적인 다양성 분석과 결합한다면 더욱 넓은 관점에서 통합적인 생물다양성과 생태 현황 평가를 수행할 수 있다 (Li et al. 2019).
최근, 대표적 응용 생태학 분야인 복원생태에서 역시 계통발생학적 다양성의 활용이 증가하고 있다 (Hipp et al. 2015, Ribeiro et al. 2018, Lishawa et al. 2019). 또한 산림 작업 기법의 적용 후 뒤따르는 다양성 변화 연구에서 역시 계통발생학적 다양성이 활용되고 있다 (Zhou et al. 2018). 계통발생은 기능 형질의 진화적 역사를 넘는 통합성을 가지며, 형질 기반 예측 도구 (예를 들면 기능 다양성)보다 뛰어난 생태적 요인에 대한 유용한 예측 도구가 되고 있다 (Cadotte et al. 2009).
다수의 생태 이론과 분석의 배경은 지위 (niche) 개념, 특히 종 간 지위 차이에서 발생하는 생태 기능의 차이가 갖는 함의이다. 그러나 다양한 종들이 차지하고 있는 지위 공간의 직접적 측정은 개념적으로도 그리고 방법론적으로 쉽지 않다. 이러한 문제의 한 부분으로서 측정하거나 평가하기 난해한 어떤 종의 지위는 해당 생물이 자라고, 살아가고, 번식할 수 있는 어떤 (또한 수많은) 조건으로 표현되는 다차원에 걸쳐 역동적으로 변화하는 것이기 때문이다 (Garnier et al. 2016). 대표적인 두 가지 접근법인 종의 “형질” (McGill et al. 2006), 그리고 “계통수에서 종 간 거리” (Webb et al. 2002)를 활용한 지위 특성 측정을 통해 이러한 어려움을 극복하고자 노력하였다. 두 가지 접근법 모두 종의 지위에 가지는 다차원성에 대해 설명할 수 있다; 형질 기반 측정은 지위 축과 관계 있는 다양한 형질 측정 값을 통합하여 표현할 수 있고 (Violle and Jiang 2009), 계통발생학적 거리는 종들 사이에 다양한 양상으로 나타나는 진화적 차이점들의 축적을 반영하고 있다. 다시 말하면, 어떤 형질을 통해 뚜렷한 환경 경사를 따르며 나타내는 종의 성장, 생존, 번식의 활성 (performance)을 측정하고자 한다면, 그 형질들은 종의 지위 정보를 제공할 수 있어야 한다 (Garnier et al. 2016). 직관적으로, 만약 이러한 형질들이 시간과 함께 꾸준하게 진화한다면, 종 간의 진화적 상관성은 형질을 사용해 얻는 것과 유사한 지위 공간의 추정치를 생산할 수 있어야 한다. 바로 이점이 형질에 기초한 기능다양성 접근법을 뛰어넘는 계통발생학적 다양성 측정 도구가 활용되고 있는 밑바탕이다 (Kraft et al. 2007).
일반적으로 가까운 계통 관계는 계통적으로 형질들을 보전하는 것을 의미하기 때문에 (Vane-Wright et al. 1991), 계통발생학적 다양성은 기능 형질 값들을 얻기 어려울 때 기능적 다양성 측정을 대신하여 사용될 수 있다 (Flynn et al. 2011, Xu et al. 2019). 그리고 α-다양성과 기능 다양성을 넘어 계통발생학적 다양성 평가는 새로운 다양성 측정과 평가, 그리고 생태계 기능과 보전학적 의미 발굴에 대한 촉매제가 되고 있다 (Cadotte et al. 2008). 계통발생학적 다양성은 앞으로 지역, 국가, 장소의 다공간 수준에서 진행되는 생물다양성 소실의 영향을 이해하고, 예측하며 그리고 잠재적 개선 효과에 대한 결정적인 정보를 제공할 수 있다 (이후승 et al. 2021). 일반적으로 생물종의 멸종 잠재성은 해당 종의 개체군 크기와 역의 상관을 나타내며, 모든 범위의 분류군의 모든 개체군 합은 보통 제한적이기 때문에, 계통발생적 널 모델에는 반드시 지역 생물상에 관한 정보가 바탕이 된다 (Hubbell 2001). 교란된 생태 군집은 유사한 속성 (계통적으로 가까운)을 가진 종들로 조합된다 (Helmus et al. 2010). 대표적으로 길가 식생을 구성하는 종들의 군집, 묵논과 묵밭 같은 신속한 천이 과정에서 관찰되는 종의 교대 과정을 좋은 예로 들 수 있다.
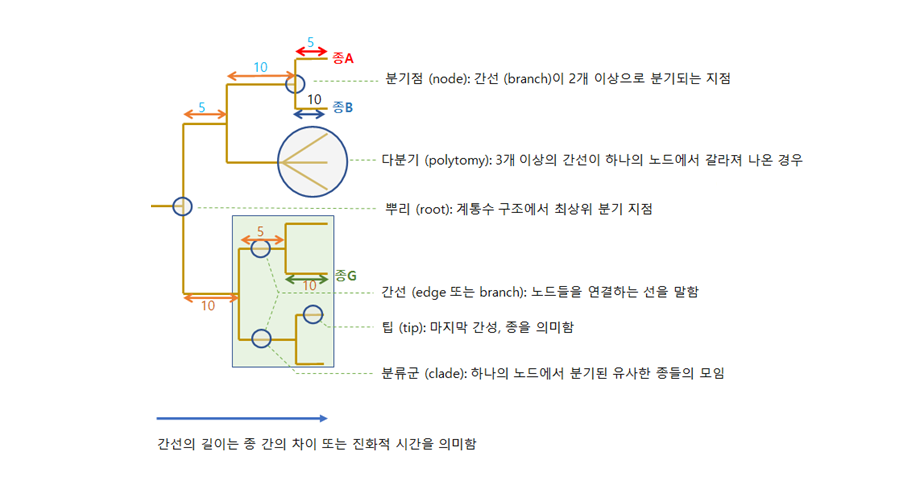
일반적으로 계통도는 아래 그림과 같이 뿌리, 분기점, 간선, 팁으로 구조화되어 있다 (Baum 2008). 뿌리와 분기점은 유사해 보일 수 있지만, 뿌리는 계통수에서 최상위 분기 지점으로서 해당 계통수에서 속하는 모든 종의 조상을 나타내며, 분기점은 뿌리 아래에서 2개 이상의 간선으로 나누어지는 지점으로서 종과 종이 구분되는 위치를 말한다. 간선은 분기점들을 연결하는 선으로서 길이의 차이에 따라 종 간의 관계성을 나타낸다. 종합적으로 종과 종 사이의 계통발생학적 거리는 종간의 차이, 그리고 진화적 차원에서의 시간 차이를 의미한다.
계통발생학적 다양성의 측정은 기능 다양성과 동일하게 간선의 길이를 활용한다. 아래 그림에서 간선을 따라 진행된 진화적 형질 변화는 간선의 길이로 구하며, 종A는 20 (5+10+5), 종B는 25 (10+10+5), 종G는25 (10+5+10)로 계산된다. 따라서 종A와 종B 사이의 계통발생학적 거리는 55로 표현된다. 만약 조사한 어떤 군집이 종A, B, G로 구성되어 있다면, 해당 군집의 계통발생학적 다양성은 (5+10+5)+(10+10+5)+(10+5+10)으로 구할 수 있다.
계통발생학적 다양성은 α 수준의 종풍부도에서 드러나지 않는 몇몇 한계를 극복할 수 있는 장점이 있다. 예를 들면, 조사된 두 지역의 종수가 동일할 때는 지역 간 차이를 추출하기 어렵지만, 계통발생학적 다양성에서 측정된 값이 클수록 계통수 내 거리가 먼 종들이 공존하며, 작은 값일수록 계통수 내 거리가 가까운 종들이 함께 살고 있는 것으로 차이를 표현할 수 있다. 이것은 앞서 언급한 특정 환경에서 종의 지위 분화 정도를 나타내는 것으로서, 기능 다양성 척도와 마찬가지로 생물다양성을 평가하는 핵심 배경이다.
Cadotte and Davies (2016), 이후승 (2021)에서 수정함
<참고자료>
Baum, D. 2008. Reading a Phylogenetic Tree: The Meaning of Monophyletic Groups. Nature Education 1:190.
Cadotte, M. W., B. J. Cardinale, and T. H. Oakley. 2008. Evolutionary history and the effect of biodiversity on plant productivity. Proceedings of National Academy of Sciences 105:17012-17017.
Cadotte, M. W., J. Cavender-Bares, D. Tilman, and T. H. Oakley. 2009. Using phylogenetic, functional and trait diversity to understand patterns of plant community productivity. PLoS One 4:e5695.
Cadotte, M. W., R. Dinnage, and D. Tilman. 2012. Phylogenetic diversity promotes ecosystem stability. Ecology 93:s223-s233.
Flynn, D. F. B., N. Mirotchnick, M. I. Palmer, and S. Naeem. 2011. Functional and phylogenetic diversity as predictors of biodiversity–ecosystem‐function relationships. Ecology 92:1573-1581.
Garnier, E., M.-L. Navas, and K. Grigulis. 2016. Plant functional diversity. Oxford University Press, Oxford, UK.
Helmus, M. R., W. B. Keller, M. J. Paterson, N. D. Yan, C. H. Cannon, and J. A. Rusak. 2010. Communities contain closely related species during ecosystem disturbance. Ecol Lett 13:162-174.
Hipp, A. L., D. J. Larkin, R. S. Barak, M. L. Bowles, M. W. Cadotte, S. K. Jacobi, E. Lonsdorf, B. C. Scharenbroch, E. Williams, and E. Weiher. 2015. Phylogeny in the service of ecological restoration. Am J Bot 102:647-648.
Hubbell, S. P. 2001. The unified neutral theory of biodiversity and biogeography (MPB-32). Princeton University Press.
Kraft, N. J. B., W. K. Cornwell, C. O. Webb, and D. D. Ackerly. 2007. Trait evolution, community assembly, and the phylogenetic structure of ecological communities. The American Naturalist 170:271-283.
Li, D., L. Trotta, H. E. Marx, J. M. Allen, M. Sun, D. E. Soltis, P. S. Soltis, R. P. Guralnick, and B. Baiser. 2019. For common community phylogenetic analyses, go ahead and use synthesis phylogenies. Ecology 100:e02788.
Lishawa, S. C., B. A. Lawrence, D. A. Albert, D. J. Larkin, and N. C. Tuchman. 2019. Invasive species removal increases species and phylogenetic diversity of wetland plant communities. Ecol Evol 9:6231-6244.
McGill, B. J., B. J. Enquist, E. Weiher, and M. Westoby. 2006. Rebuilding community ecology from functional traits. Trends in Ecology & Evolution 21:178-185.
Ribeiro, R. A., T. C. Giannini, M. Gastauer, M. Awade, and J. O. Siqueira. 2018. Topsoil application during the rehabilitation of a manganese tailing dam increases plant taxonomic, phylogenetic and functional diversity. J Environ Manage 227:386-394.
Srivastava, D. S., M. W. Cadotte, A. A. MacDonald, R. G. Marushia, and N. Mirotchnick. 2012. Phylogenetic diversity and the functioning of ecosystems. Ecol Lett 15:637-648.
Vane-Wright, R. I., C. J. Humphries, and P. H. Williams. 1991. What to protect?-Systematics and the agony of choice. Biological Conservation 55:235-254.
Violle, C., and L. Jiang. 2009. Towards a trait-based quantification of species niche. Journal of Plant Ecology 2:87-93.
Webb, C. O., D. D. Ackerly, M. A. McPeek, and M. J. Donoghue. 2002. Phylogenies and Community Ecology. Annual Review of Ecology and Systematics 33:475-505.
Xu, J., H. Dang, M. Wang, Y. Chai, Y. Guo, Y. Chen, C. Zhang, and M. Yue. 2019. Is Phylogeny More Useful than Functional Traits for Assessing Diversity Patterns Under Community Assembly Processes? Forests 10.
Zhou, X., H. Zhu, Y. Wen, U. M. Goodale, X. Li, Y. You, D. Ye, and H. Liang. 2018. Effects of understory management on trade-offs and synergies between biomass carbon stock, plant diversity and timber production in eucalyptus plantations. Forest Ecology and Management 410:164-173.
이후승, 정슬기, 주용준, 사공희, and 지민규. 2021. 생물다양성협약 대응을 위한 계통적 다양성 평가의 정책적 활용방안. 환경포럼 25.